Please put an active hyperlink to our site (www.rusnature.info) when you copy the materials from this page
Biomes and Regions of Northern Eurasia
The Mountains of Central Asia and Kazakhstan
<<< Modern Glaciation | Biomes & Regions Index | Mountains and
People: Past, Present, and Future >>>
Mountainous Biota
The mountains of Central Asia and Kazakhstan are one of the four centres of biological
diversity within the territory of the FSU. The peculiarity of biota is due to its mixed
character: Indo-Hymalayan, Mongolian, Eurasian, and Mediterranean species are represented
as well as local endemics. Under equal conditions, species-richness and endemism in the
mountains are always higher than on the surrounding plains, first, because mountains serve
as refugia during unfavourable climatic epochs, and second, because of higher habitat
diversity. Thus, the flora of the Central Asian mountains comprises about 5500 species,
while the surrounding deserts accommodate about 2000 species (Table 16.6).
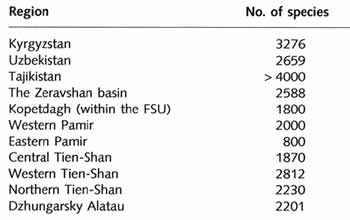
Table 16.6 Number of plant species in various regions of the Central
Asian mountains
Altitudinal vegetation zones range from the deserts of the foothills to the alpine
environments of the high mountains (Figure 16.8).
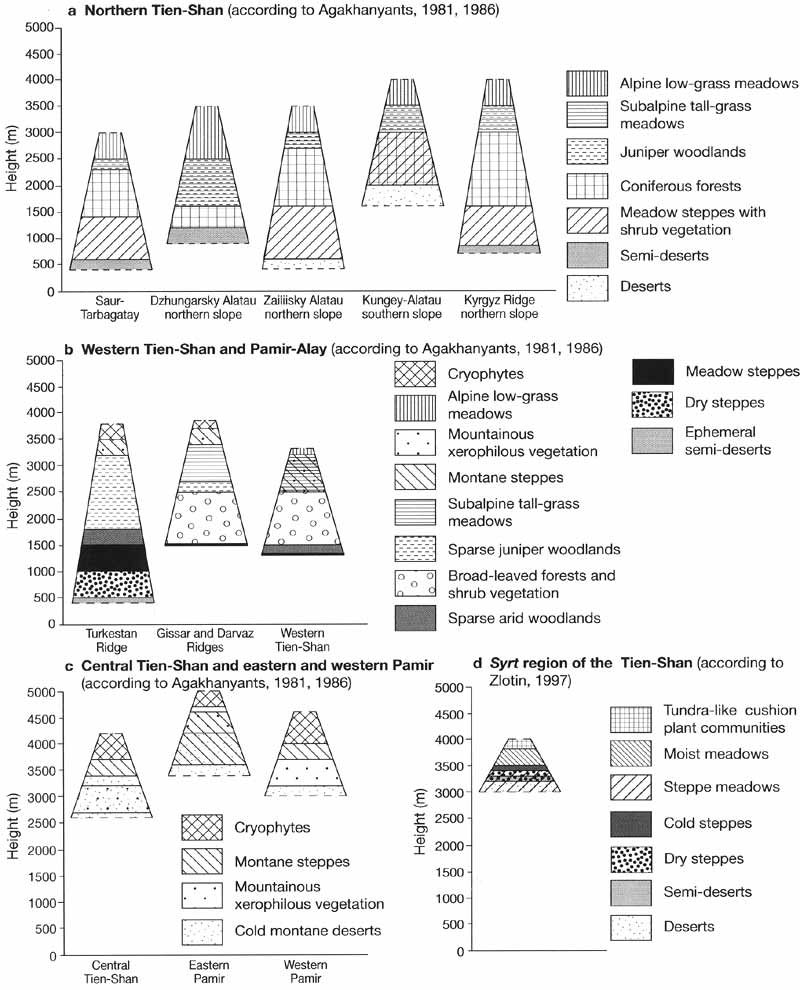
Fig. 16.8 Altitudinal zonation of vegetation.
There are two general types of altitudinal sequences: a more humid sequence with forest
belts, which occurs in the Dzungarsky Alatau and in the northern part of the Tien-Shan
mainly on the slopes of northern and western exposure, and an arid sequence typical of
more southern locations (e.g., the south-western Tien-Shan, Pamir-Alay) and slopes located
in the rainshadow in which forests are not represented as a continuous belt. Both types of
altitudinal zonation are most clearly expressed in the marginal regions. In the arid
interior regions (i.e., the central Tien-Shan and eastern Pamir), the occurrence of
elevated plateaux and continentality of climate mask altitudinal differentiation. Human
activities, which began in the Central Asian mountains thousands of years ago, have had
strong impacts on vegetation, ranging from creating areas with a rich collection of
cultivated plants to the destruction of montane forests. In this chapter, most attention
is given to the sequences with forest belts and for the Pamir and Tien-Shan these are
based on the models by Agakhanyants (1981, 1986). Stanyukovich (1973), Walter and Box
(1983a, b), and Ogureeva (1999) provide more comprehensive surveys.
Kopetdagh
Biogeographers are in agreement that the Kopetdagh (together with the rest of a larger
Turkmen-Khorossan mountainous system, the major part of which is located in Iran) is a
separate biogeographic entity. However, because of the transitional nature of the biota,
its status and affiliation with units of the higher level are debated (V. Fet, 1994). The
most distinct feature of the Kopetdagh biota is the combination of Iranian and Turanian
species, which constitute the core of its flora, with European and Mediterranean species
within a very distinctly defined geographical area. For many species, the Kopetdagh marks
a limit of their distribution. The specific biogeographic position, combined with the
prolonged isolatioji in the Paleogene-early Neogene, resulted in the development of
numerous endemics. Many species have a very narrow range of distribution which
distinguishes this area from the conventional transitional zones. Plant endemism is
unusually high: about 18 per cent of plants found in the Kopetdagh are endemic (Kamelin,
1970). Most of them exhibit a taxonomic relationship to the Iranian and eastern
Mediterranean species, indicating that the vegetation of the Kopetdagh evolved from the
ancient Mediterranean flora under the influence of two major factors: continuous
aridization since the end of the Tertiary and orographic uplift (Kamelin, 1973; Kurbanov,
1994). In the Tertiary, the south of Central Asia was dominated by subtropical evergreen
forests of the laurel type and by tall-grass savannahs. However, even arid woodlands,
which had been formed in the Kopetdagh by the middle Pliocene, became impoverished later
and a facilitated evolution of the xerophilous flora occurred. There are a number of texts
dealing with the history of the Kopetdagh biota (Kamelin, 1973; Korovin, 1961;
Ovchinnikov, 1971) and a more recent review has been given by Kurbanov (1994).
Although the mountains do not reach the snow line and only in the central Kopetdagh
subalpine plant communities develop between 2500 m and 3000 m, vertical zonality has a
complex structure (G. H. Fet, 1994). One of the first models of altitudinal zonation was
proposed by Chernyakhovskaya (1927) who distinguished six vegetation zones: submontane
plain, sagebrush steppe, grass and herbaceous-grass steppe, juniper forests, highland
vegetation, canyon vegetation, and intrazonal vegetation of rocky outcrops. All subsequent
schemes were largely variations of this original model. However, because boundaries
between the altitudinal belts are not distinct and plant communities have a mosaic
distribution and vegetation is often disturbed, there is still a disagreement on
altitudinal division (as well as regionalization) of the Kopetdagh vegetation. Thus,
Kamelin (1970), who also distinguishes six altitudinal zones, comments that only two are
distinctly different: a shiblyak (a diverse community of low tree and shrub species) and
semi-savannah belt between 400 m and 1500 m and a juniper and steppe belt between 1600 m
and 2800 m.
In the foothills, including adyrs which occupy the area between 400 m and 800 m,
subtropical desert vegetation develops. Sagebrush communities predominate together with
numerous ephemerous and ephemeroid species whose participation predetermines a clearly
expressed seasonal change in vegetation. A large extent of soil erosion, associated with
poor vegetation cover and strongly enhanced by overgrazing, leads to the widespread
occurrence of species adapted to stony soils. Under the conditions of heavy grazing,
sagebrush coverage declines from 40 per cent to 5-10 per cent and in the strongly eroded
locations petrophytes may completely replace sagebrush communities (G. H. Fet, 1994). The
low mountain belt, between 800-900 m and 1200-1400 m, is occupied by the tall-grass
semi-savannah composed mainly of the communities dominated by the perennial grass
Elytrigia trichophora. Ephemeroid species and meadow steppe herbaceous vegetation
co-dominate in the lower part of this belt while in the upper part the participation of
the steppe species, such as Festuca valesiaca, increases. At higher altitudes, between
1200-1400 m and 1900-2300 m, steppe vegetation develops, dominated by Stipa spp. and
Festuca valesiaca. Above this boundary, upland xerophilous plants predominate.
Although woody vegetation occupies small areas in the Kopetdagh, developing mainly on
the northern slopes, in river valleys and along the beds of seasonal streams, tree and
shrub communities are diverse. Most widespread are shiblyak communities, dominated by
Paliurus spina-christi, Acer turcomanicum, Primus, Amygdalus, and Rosa spp. (G. H. Fet,
1994; Popov, 1994). Shiblyak comprises a variety of fruit trees and vines suitable for
cultivation, including pomegranate, fig, wild grapes, apple, cherry, and many others. It
develops mainly in the low mountains: Paliurus spina-christi occurs between 400 m and 800
m, Acer turcomanicum is limited to the 800-1500 m zone and only Berberis integerrima and
Crataegus pontica can penetrate the middle mountains to an altitude of 1900 m. Typical of
the higher altitudes are juniper woodlands. These are composed ofjuniperus turcornanica,
the only juniper species found in the Kopetdagh and one of the most drought-resistant
species which can grow under the conditions of annual precipitation below 150 mm and
summer temperature above 46°C (Popov, 1994). While at present juniper woodlands are
confined mainly to the middle mountains, as recently as in the 19th century they were
widespread at 400-600 m. Juniper, which was widely used as firewood, is also the only tree
in the Kopetdagh which can provide timber for construction. Between the 1930s and the
1960s, the area occupied by juniper woodlands declined from an estimated 79 000 ha to 20
000 ha (Koksharova, 1970). The easily accessible lower mountains in the western ridges
were affected particularly badly and only on the steep slopes of the central ridges does
tree cover reach 40-50 per cent and old trees reaching 18-20 m in height can still be
found (Popov, 1994).
Riparian forests develop in the south-western Kopetdagh in the mountain river valleys
which have a more humid climate as a result of their proximity to the Caspian. Such
forests, composed mainly of Juglans regia with the participation of Fraxinus syriaca,
Ulmus carpinifolia, Thelycrania meyeri, and Platanus orientalis are confined to a very
narrow (between 50 m and 100 m) zone along rivers at altitudes from 1000 m to 1500 m.
Vines, which are not found in the Central Asian mountains further east, are common and a
representative of the relatively moisture-demanding Miocene flora, Periploca gracea,
occurs (Kamelin and Zabelina, 1987). Because of the abundance of fruit trees and vines,
riparian forests of the south-western Kopetdagh have been a centre of genetic and
selection studies conducted by the Vavilov Institute of Plant Breeding since the 1930s. Of
particular practical interest was such a valuable species as Juglans regia and already in
the 1930s, a lack of young trees was observed resulting from overharvesting of walnut
seeds (Popov, 1994). By the early 1990s, the population of Juglans regia had been reduced
to less than 2000 trees partly due to the continuing overharvesting and partly due to the
occurrence of unusually strong mudflows in the 1980s. With the continuing depletion of
Juglans regia, the participation of light-loving shrubs and trees increases. In
particular, a hydrophilous relict species of Mediterranean origin, Platanus orientalis,
which in Central Asia occurs only in riparian habitats, has become more widespread. The
presence of relatively moisture-demanding riparian forests in the southwestern Kopetdagh
and their composition as well as the proximity to the Transcaucasia, led to suggestions
that the flora and fauna of the Kopetdagh belong to the Hyrcanian type. Hyrcania, which
comprises the southern and south-western shores of the Caspian Sea, is a unique relict
biogeographical region. As V. Fet (1994) points out, there are indeed many relict elements
in the riparian biota of the south-western Kopetdagh. Some specific Hyrcanian elements
(such as the ground beetle Broscus karelini and the darkling beetle Metadisa virdis) can
be found in the humid valleys of the Kopetdagh and many genera have variant species (e.g.,
the spiders Dysdera concinna in Hyrcania and D. pococki in the Kopetdagh). However, the
Tertiary relicts, typical of the Kopetdagh riparian forests, are characteristic of the
whole of the ancient Mediterranean region. Species with the Hyrcanian-Kopetdagh
distribution are few, and such characteristic dominants of the Hyrcanian forests as
Quercus castaneifolia and Parrotia persica do not occur in the Kopetdagh. Therefore,
assigning the Kopetdagh to the Hyrcanian biogeographical region is not justified but
questions remain about the ways of penetrations of the European, Mediterranean, and
Caucasian elements eastwards which, being limited by the Caspian in the north and deserts
of Iran in the south, led through Hyrcania.
Tien-Shan and Pamir
The enormous heights of the Tien-Shan and Pamir predetermine the development of full
spectrum of altitudinal zones. However, because of the complex orography and
non-uniformity of climate, floristic variations occur between ridges and slopes of
different exposure. Acknowledging the strong heterogeneity of the vegetation, Agakhanyants
(1981) introduced a term 'mountain plakor' and suggested that models of vertical
zona-tions can only be developed for these 'typical' slopes. 'Mountain plakor' is
certainly a contradiction in terms, since in the Russian-language literature the term
'plakor' is used to describe flat surfaces or watersheds on which zonal vegetation
develops, but its use certainly highlights the complexity of vegetation classification.
Agakhanyants (1981, 1986) distinguishes six biogeographical provinces according to the
history of floristic development, modern composition of vegetation and vertical structure
of biota: northern Tien-Shan, south-western Tien-Shan, western Pamir-Alay, western Pamir,
and central Tien-Shan and eastern Pamir (Figure 16.1), although Zlotin (1975) argues that
because of different altitidues, degree of continentality, and specific features of
community structures, central Tien-Shan and eastern Pamir should not be viewed as a single
province.
Northern Tien-Shan
The northern Tien-Shan (including Dzhungarsky Alatau, Saur, Tarbagatay, and Zailiisky,
Kyrgyz, and Kungey-Alatau Ridges) has a full spectrum of altitudinal zones (Figure 16.8a).
Temperate semi-deserts dominated by sagebrush communities occur in the foothills extending
from the westernmost regions of the Tien-Shan to the Dzhungarsky Alatau. Above these, a
zone of feathergrass steppe develops typical of which is a widespread occurrence of
shrubs, in particular Rosa spp. and Lonicera microphylla, on wetter and shaded slopes and
in the valleys. At the upper part of the belt, feather-grass steppes pass into steppes and
woodlands composed of Primus racemosa and Mains sieversii become a notable part of the
landscape. In contrast to the meadow steppes of the southern regions, the meadow steppes
of the northern Tien-Shan do not experience a severe deficiency in moisture and remain
green throughout the summer. The origin of the flora is also different: while many plants
in the southern regions are of Mediterranean origin, the flora of the meadow steppes of
the northern Tien-Shan is closely connected to the steppes of the Kazakh plains. In the
Dzhungarsky Alatau, species of the Altay-Siberian origin predominate while for many of the
Central Asian species the ridge marks the northern and eastern boundaries of their
distribution. Forests succeed steppes first on northern slopes at an altitude of 1500-1700
m and then on southern slopes. In contrast to other regions where walnut forests prevail,
deciduous woodlands in the northern Tien-Shan, developing in valleys in the lower part of
the forest belt, are composed of Populus tremula. Characteristic of the northern Tien-Shan
are coniferous forests, comprised of Picea schrenkiana and Abies semenovii at higher
altitudes with the participation of Betula, Sorbus, and numerous shrubs in the
undergrowth. In the Dzhungarsky Alatau, Picea obovata is a typical forest-forming species.
Although this belt receives about 800 mm of precipitation per annum and is generally
reminiscent of the taiga landscapes, the dark forest soils which develop under the montane
forests have much higher humic content and lower acidity than podzols. This is because the
chemical composition of needles of Picea schrenkiana is different from that of other
species of spruce: the content of calcium oxide in needles of Picea schrenkiana reaches 44
per cent while in needles of the other spruce species it does not exceed 12 per cent
(Glazovskaya, 1955). Forests are succeeded by the floristically rich subalpine zone which
is a complex of tall-grass meadows and woodlands formed by Juniperus sibirica. Junipers
have a bush-like form at lower levels and take a creeping form at higher altitudes.
Tall-grass meadow communities are dominated by sedges (mainly Helictotrichon asiaticum,
Alopecurus songoricus, and Festuca rubra) with the participation of cold-tolerant
flowering species (e.g., Geranium spp., Anemone narcissiflora, Aster alpinus) and form
vast and productive summer pastures. At an altitude of 2800-3000 m, the alpine low-grass
meadows dominated by Kobresia spp. develop. The diversity of species as well as
productivity of the alpine meadows is considerably lower than those of the tall-grass
meadows and locally vegetation does not form a continuous cover.
Western Tien-Shan and Pamir-Alay
With respect to biogeography, there is much similarity between the south-western
Tien-Shan and the western Pamir-Alay. The western Tien-Shan has a temperate climate but
south of the Chatkalsky Ridge, climate and landscapes assume subtropical features. The
amount of precipitation received by individual ridges and slopes differ widely across the
region. The mountains of southern Tajikistan, for example, have an arid climate while the
Gissar and Darvaz receive an ample moisture supply. The uneven distribution of
precipitation predetermines local differences in altitudinal zonation particularly with
respect to the occurrence of forests. Forests develop mainly on wetter northern and
western slopes. Thus, juniper woodlands occur on the northern macroslope of the Turkestan
Ridge while its southern slope is devoid of woody vegetation and its landscape is that of
an extremely arid environment. In all, there are three types of zonal woodlands (sparse
arid Pistacia woodlands, broad-leaved deciduous woodlands, and open juniper woodlands) and
seldom are all three types represented in the altitudinal sequences.
A model of altitudinal zonation suggested by Agakhanyants (1981, 1986) for moister
slopes is shown in Figure 16.8b. The sequence begins with the belt of deserts or
semi-deserts which assume a subtropical character, expressed in the domination of
ephemeral species, in the Fergana valley. These are succeeded by dry steppe
(semi-savannah) belt, whose vegetation is composed mainly of tall forbs. Meadow steppe
dominated by large umbellates, among which Prangos pabularia and species of Ferula are
most widespread, develops above. In contrast to the northern Tien-Shan, most plants end
their growth by late July and meadow vegetation dies with increasing moisture deficit.
Under the meadows on fine alluvium, mountainous chernozem, and chernozem-like soils form
with a high humic content and a well-expressed carbonate layer. In the lower part of this
zone, shrub vegetation develops in shaded sites. On western slopes and in the peripheral
parts of the region, particularly in the Gissar, Darvaz, and Peter the First Ridges,
deciduous woodlands occur as isolated groves on wetter slopes. The main forest-forming
species in broad-leaved woodlands are Juglans regia and J. fallax with co-domination of
Acer spp. or Malus sieversii. The Fergana Ridge marks the eastern limit of distribution of
walnut groves and while they occur as far north as the Talas-Alatau, in more northern
regions walnut groves usually prefer sites protected from the north and exposed to the
south-west. In all, there are over a hundred tree and shrub species in deciduous
woodlands; fruit trees are particularly abundant but notable is the absence of oak.
Because deciduous woodlands have a mosaic distribution, their classification into a
separate altitudinal belt is debated. Thus Agakhanyants (1981, 1986) distinguishes a belt
of 'woodlands and shrubs' rather than woodlands and in the model put forward by Pavlov
(1980) woodlands are not recognized as a separate entity. In the upper part of the meadow
steppe zone, deciduous woodlands are succeeded by open juniper woodlands comprised of
Juniperus species of the Central Asian origin (J. seravschanica, J. semiglobosa, and J.
turkestanica). In more arid locations, juniper woodlands (locally with Pistacia in the
undergrowth) develop in place of broad-leaved forests and mountainous feather-grass steppe
communities occur instead of the meadow steppe vegetation. From an altitude of 2500-2700
m, woody vegetation disappears, giving way to the tall-grass meadows. Similar to the
meadow steppe zone, there is a notable domination of umbellates while in drier localities
elements of steppe vegetation occur. Chernozem-like soils develop under the subalpine
meadows but in contrast to the soils of the meadow steppe, these are devoid of carbonates
and have a brown sod layer. The subalpine meadows form the most important summer pastures
with productivity of fodder species of 0.8-1.8 tonne ha-1a-1
(Ovchinnikov, 1977). Low-grass alpine meadows succeed the tall-grass meadows and
vegetation adapted to low temperatures develops above.
Western Pamir
The western Pamir is distinguished from the western Tien-Shan and Pamir-Alay by the
absence of zonal woody vegetation. With the increasing continentality of climate, the
sequence of altitudinal zones becomes more similar to those of the eastern Pamir and
central Tien-Shan. Attempting a general model, intended for both the western and eastern
Pamir and Central Tien-Shan, Agakhanyants (1981, 1986) divided the mountains into four
belts (Figure 16.8c). What distinguishes montane deserts of the western Pamir from those
located landwards is the presence of ephemerous plants at lower altitudes. Vegetation
communities are dominated by Artemisia vachanica, A. lemanii, Hammada vachanica, Peganum
harmala, and Poa bulbosa. Xerophilous vegetation (a very broad term favoured by
Agakhanyants but not by other authors), represented mainly by the communities of
Acanthophyllum pungens, Acantholimon, Astragalus, and Onobrychis spp., develops above on
the rocky slopes. It is succeeded by feather-grass steppes. However, steppe communities
occur widely in the neighbouring zones and the altitudinal boundaries of their
distribution vary widely across the region.
Eastern Pamir and Central Tien-Shan
Characteristic of the eastern Pamir and central Tien-Shan are extremely severe
climates, with respect to both temperature and aridity, and the widespread occurrence of
permafrost above 3000 m. The duration of growing season is short and ranges from six
months at 3000 m to six weeks at 4000 m (Zlotin, 1997). The mean annual temperature is
below zero and there are no periods without frosts. Snow cover stays from two to six
months and, under the conditions of very low precipitation and strong winds, particularly
in the eastern Pamir, it is thin and uneven which causes the soil to freeze deeply. The
environments of the eastern Pamir and central Tien-Shan are similar to those of the Arctic
tundra and, as a result of these severe conditions as well as past glacia-tions,
vegetation is poor in comparison with that of the other regions of the Central Asian
mountains. Thus, in the high mountain deserts of the eastern Pamir, vegetation cover is
8-15 per cent and as few as 3-7 species are found in 100 m2 (Walter and Box,
19836). Most of the eastern Pamir and central Tien-Shan belongs to the alpine zone.
However, because of the dryness of climate, alpine vegetation develops only in better
moistened locations while most of the area is dominated by cold mountainous deserts and
steppes similar to those of Mongolia and Tibet. Due to the characteristic local levelness,
altitudinal zonality is not clearly expressed. The belt of xerophilous vegetation of rocky
deserts is fragmented in the Tien-Shan and the belt of mountain steppes has a patchy
character in the eastern Pamir.
Focusing on the central Tien-Shan, Zlotin (1997) suggested a more detailed model which
classifies vegetation into seven altitudinal 'steps' within the alpine belt (Figure
16.8d). This scheme recognizes the alpine meadows as a separate altitudinal step because
in the syrt regions of the Tien-Shan meadow communities may occupy over 80 per cent of the
area, reflecting the relatively high moisture availability. Meadow communities are
dominated by species of Kobresia and in the Tien-Shan they include about 70-80 species of
flowering plants (Zlotin, 1997). The humic content of soils developing under the Kobresia
meadows reaches 8-11 per cent. On a broader scale, meadows are less notable as a separate
step. Outside the syrt region, meadows occupy not more than 6-8 per cent of the territory,
forming intrazonal communities in stream valleys, while steppe and desert environments
account for about 50 per cent and 20 per cent, respectively.
According to the scheme put forward by Agakhanyants (1981, 1986), cold deserts occupy
the lowest belt (in the central Tien-Shan, semi-deserts, dominated mainly by Artemisia,
develop in the intermontane depressions between 1500 m and 2500 m with elements of dry
steppe feather-grass vegetation in the localities with better moisture supply). On gently
sloping surfaces, Artemisia rhodantha, Ajania tibetica, Kalidium screnkianum, and Limonium
hoeltzeri form mixed communities. Typical of drier locations, especially in the eastern
Pamir, is a dwarf shrub Eurotia ceratoides — the only local plant which can be used as
fuel wood. On the steeper, rockier slopes, xerophilous vegetation develops, formed by
Onobrychis echidna, Hedysarum minjanense, Acantholimon, and Astragalus spp. One of the
plant adaptations to the severe conditions is their cushion-like shape. Low temperatures
and strong winds set a limit to the vertical growth of plants, which expand in a
horizontal direction, and whereas fine soil is blown on to the edges of the plant, it
develops a characteristic circular form creating within itself a specific moisture and
temperature regime. Large cushions often retain seeds of other plants transported by wind
and the intergrowth of one plant with another is common. Deserts and xerophilous
vegetation are succeeded by montane steppes, formed mainly by species of Stipa, Festuca,
and Helictrotrichon, which are particularly typical of the syrt regions of the Tien-Shan.
Above, vegetation adapted to low temperatures occurs in place of alpine meadows. In the
lower part of the belt, cushion-plant communities similar to those of the frost-boil
Arctic tundra develop. In the central Tien-Shan, they are dominated by large (about 50 cm
in diameter) cushions ofDryadanthe tetrandra which, covering between 10 per cent and 30
per cent of the surface, constitutes about 9 0 per cent of the total plant reserve
(Zlotin, 1997). Typical of the eastern Pamir are cushions of Acantholimon. Herb
communities, similar to those of the polar deserts, occur at the uppermost limit of
vegetation where higher plants do not form a continuous cover.
<<< Modern Glaciation | Biomes & Regions Index | Mountains and
People: Past, Present, and Future >>>
Contents of the Mountains of Central Asia and
Kazakhstan section:
Other sections of Biomes & Regions:
|
|







